Ecology ∩ Statistics ∩ Data Science

Welcome! I am a data scientist at Getty Images where I work on many fun projects, including the search trends tool VisualGPS Insights.
I also do research in ecology and was formerly a postdoctoral researcher at the University of British Columbia and University of North Carolina at Chapel Hill.
Broadly, I am interested in developing accurate and/or useful models of biological systems, especially systems that are stochastic, multivariate, and that change over time.
Recent research has focused on range expansion, methods for estimating fish catch, and insect life history. Read more here.
You can download my CV here.
You can email me here.
News
-
September 2024: Paper on shifts in phenology across trophic levels accepted by Nature Ecology & Evolution
-
July 2024: Paper on estimating phylogenetic constraints on phenological responses out accepted by Nature Climate Change
-
April 2024: Paper on joint estimation of angler effort on Lake Nipissing out in Fisheries Research
Research
Range shifts
The geographic ranges of many species are shifting as a result of climate change and human introductions. I use modelling to understand precisely how these shifts occur and what, if any, interventions can stop or slow them.
Previous work (Legault et al. 2020) found that competitive interactions affect both the speed and shape of range expansion.
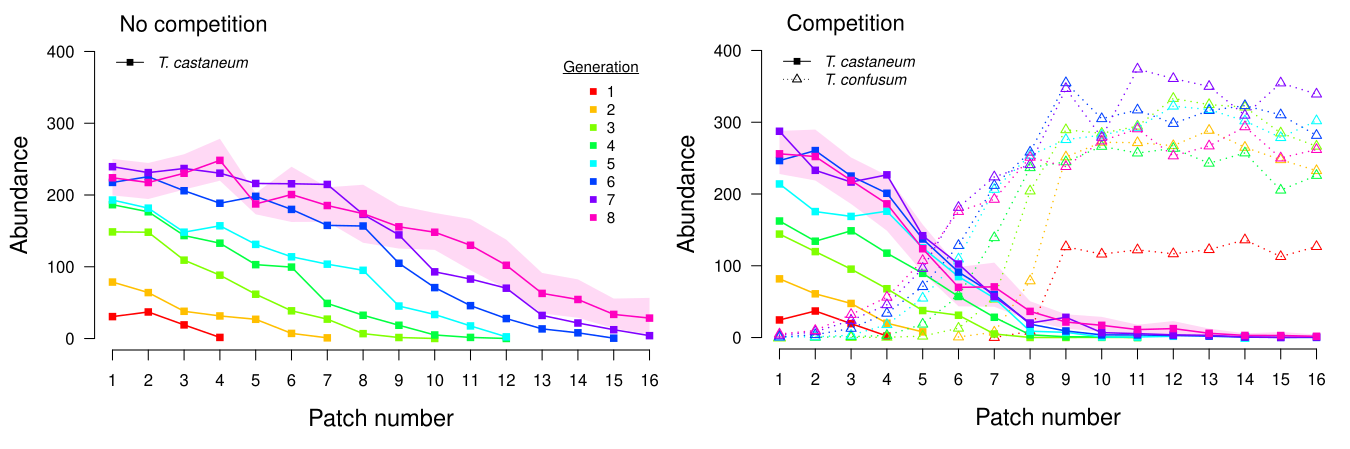
Experimental data showing the impact of interspecific competition on range expansion across 8 generations. Lines are mean abundances across 16 patch landscapes (15 replicates). Shaded areas are 95% confidence intervals for generation 8.
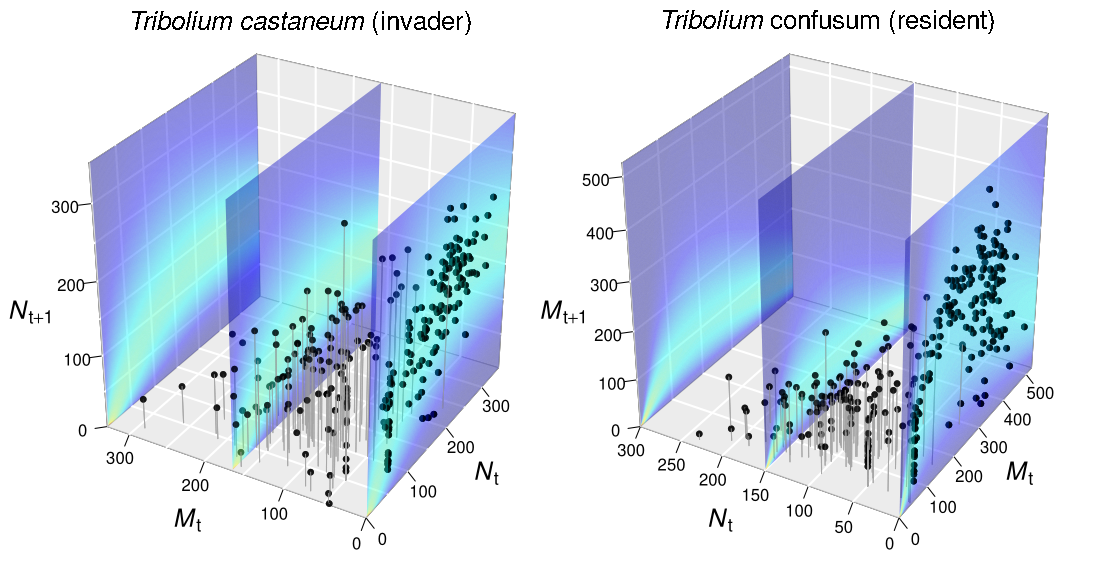
Fit of a stochastic growth model for Tribolium castaneum (left) and Tribolium confusum (right). Points are experimental data and slices show parts of the three-dimensional probability volume of the fitted model (warmer colors = higher probability). N is abundance of T. castaneum and M is abundance of T. confusum.
Improved estimation of fish catch
I am currently collaborating with researchers at the Ontario Ministry of Natural Resources and Forestry (Aquatic Research and Monitoring Section) to develop more accurate and more efficient approaches for estimating angler effort and catch in winter fisheries.
For example, Tucker et al. 2024 uses joint estimation to combine data from multiple types of creel surveys taken on Lake Nipissing.
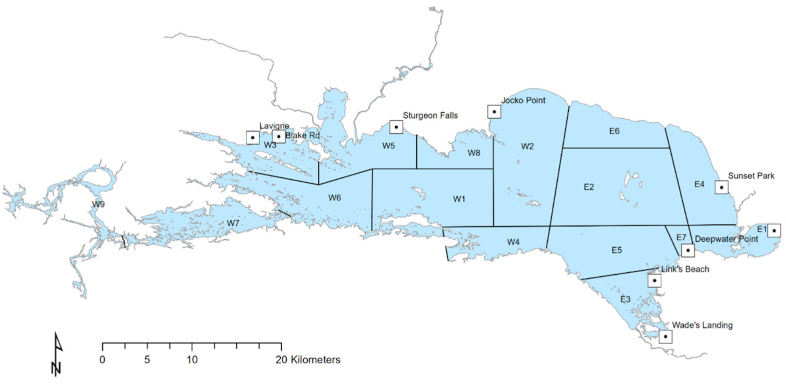
Map of Lake Nipissing in Ontario, with the locations of the nine access creel sites that were surveyed in the winter of 2023
Life history trade-offs
Life history traits (e.g., maturation time, mass at maturity) frequently co-vary and exhibit important trade-offs. Characterizing such trade-offs is necessary for predicting lifetime fitness and optimal evolutionary strategies. I develop and validate multivariate life history models (Legault and Kingsolver 2020, Parker et al. 2023), in order to better understand how these trade-offs manifest in natural systems.
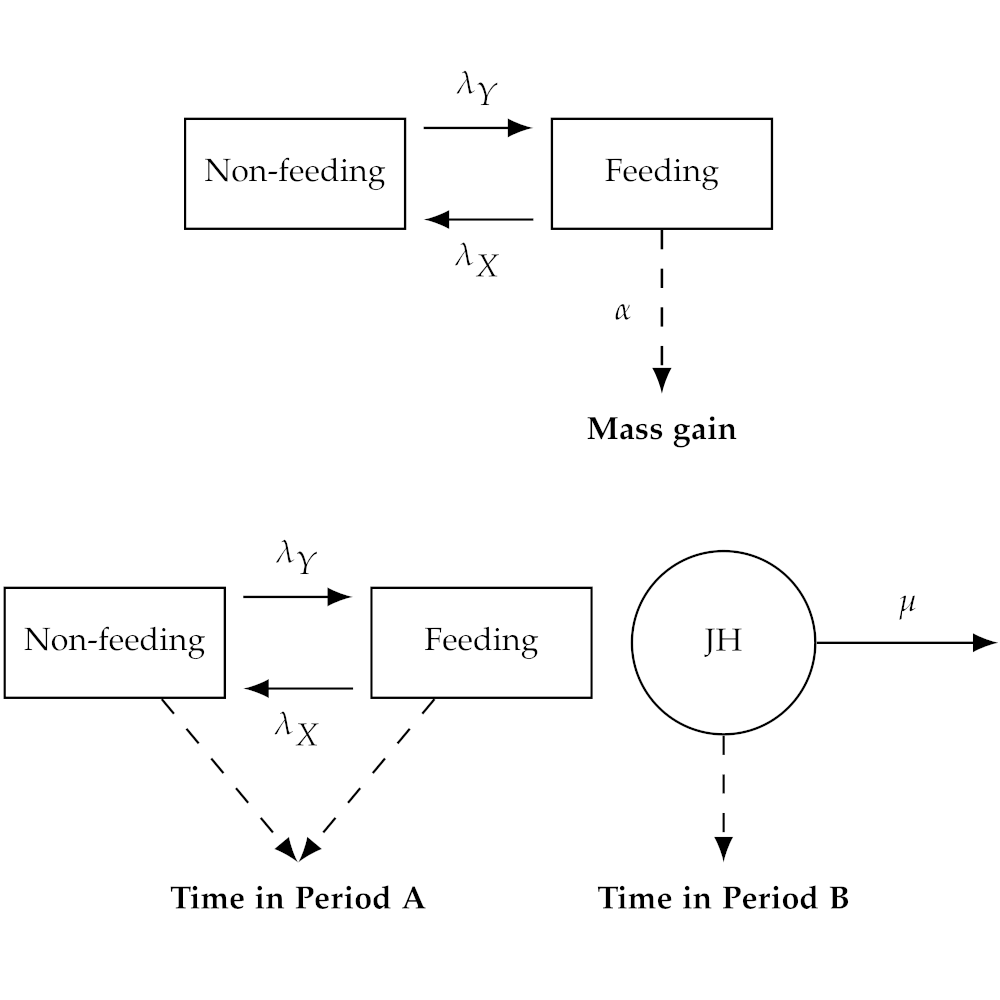
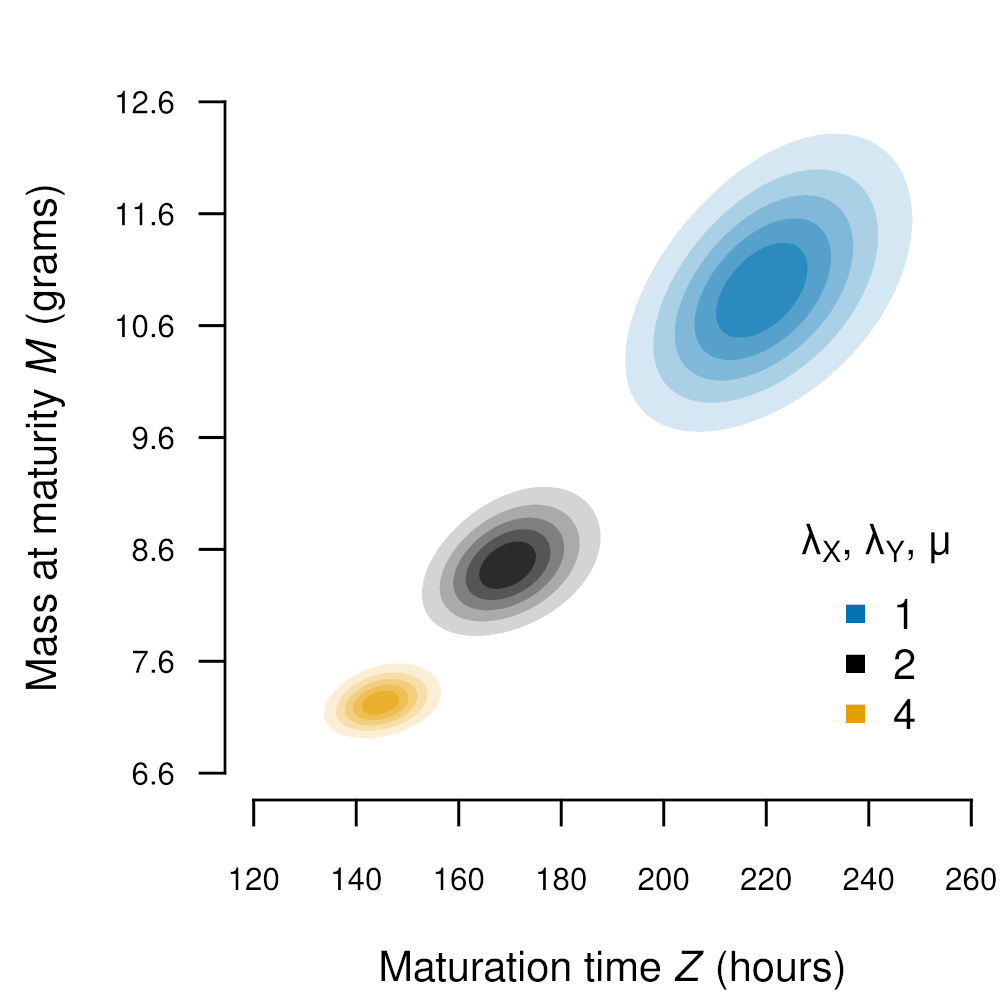
Diagram and associated contour plot of a multivariate life history model of Manduca sexta development.
Resources
![]()
Legault et al. 2020: Data and R Scripts for reproducing the analysis of range expansion. Click here to download the repository.
Legault and Kingsolver 2020: R Scripts for simulating stochastic model of insect development. Click here to download the repository.

Legault et al. 2019: R Scripts for simulating stochastic consumer-resource model. Click here to download the repository
Links
-
I wrote a parody song about a fabled land called R1 Institution. Hope you enjoy!
-
Univariate Distribution Relationships: Interactive chart of 76 univariate distributions
-
NIST Digital Library of Mathematical Functions: Comprehensive reference on special mathematical functions
-
On-line Encyclopedia of Integer Sequences (OEIS): Information on useful number sequences (e.g., A092677, a sequence used for evaluating the inverse error function)
-
mkdocs: Quick and easy static site generator using
pythonandmarkdown -
Iosevka: An open source typeface with many styles and ligature sets. Great for coding.